
Charles Darwin
Imagino que muchos de los lectores que se enfrenten a este artículo con cierta curiosidad o llamados por lo confuso del título me tildarán de “loco”. Antes de que lo hagan les diré que lo que aquí les planteo es una crítica al pensamiento darwinista clásico. No porque sea falaz (todo lo contrario), sino porque puede resultar incompleto si lo vemos a la luz de unos organismos muy especiales como son la fauna de Burgess Shale. Con este nombre se conoce a un grupo de organismos muy diversos y que se desarrollaron según se estima en un corto período de tiempo (entendiéndose siempre en la escala de tiempo geológico).
El modelo del llenado del barrido ecológico podría explicarse perfectamente reduciendo al máximo la teoría darwiniana convencional, recogiendo la obra del naturalista británico en la siguiente afirmación: “El organismo propone y el ambiente dispone”. Como es sabido, los organismos proporcionan la materia prima en forma de variación genética (que se acaba expresando finalmente en diferencias morfológicas constatables por nosotros, humildes intérpretes del proceso evolutivo). El cambio evolutivo se produce pues gracias a la selección natural (fuerza impulsora que rige el proceso y que podemos decir que “surgen” del ambiente circundante, tanto físico como de la interacción con otros organismos).
Si a continuación nos preguntamos acerca de la singularidad ambiental que puede engendrar tal disparidad anatómica y funcional como la que tuvo lugar en el Cámbrico, nos asalta una respuesta evidente. Y es que en este período lo que se produjo fue el primer llenado de un barril ecológico, siendo ésta una época de oportunidades sin parangón, de tal forma que casi cualquier forma de vida (por rocambolesca que pueda resultar) podía encontrar su lugar. He ahí que la vida radiara al espacio vacío proliferando a tasas logarítmicas. En este bullicio, la experimentación reinaba en un mundo prácticamente libre de competencia por primera vez, puesto que apenas se conoce fauna anterior a la de Burgess Shale.

Fósil de Wiwaxia. Este ejemplar se encuentra en el museo de la Smithsoniana.
En teoría darwiniana, la competencia es el gran regulador. Darwin concebía el mundo metafóricamente como un tronco con muchas cuñas (que representaría a las especies) apretadamente clavadas. Así, una nueva especie únicamente puede entrar en este mundo asestando un golpe en alguna rendija y haciendo saltar otra cuña previamente establecida, pasando a ocupar su lugar (rol ecológico). Así, la diversidad se autorregularía y de esta forma, mientras la explosión del Cámbrico iba llenando este barril iba imponiendo una condición: “Todo cambio posterior tendría lugar mediante un proceso más lento de competencia y sustitución”. Pero esta perspectiva darwiniana plantea de manera concomitante la objeción obvia a este modelo, pues la vida ha sufrido sorprendentes extinciones en masa (la del Pérmico pudo haber acabado con el 95% o más de todas las especies existentes), y hasta le fecha el fenómeno de disparidad acaecido en Burgess Shale nunca ha vuelto a ocurrir. La vida se rediversificó después de este cataclismo, pero no surgieron nuevos filos. Los recolonizadores de esta Tierra depauperada se mantuvieron dentro de las restricciones de unos pocos diseños anatómicos previos (actualmente no existen organismos con la anatomía de Wiwaxia, Hallucigenia ni Opabinia por citar a algunos). ¿Qué puede haber provocado esta situación?
Muchos autores defienden que esta radiación sin precedentes no puede explicarse sólo por la ecología del medio externo en que se desarrollaban estos organismos. No aceptan que si los organismos siempre tienen el potencial para poder diversificarse de esta manera, nunca, ni siquiera una vez haya surgido un nuevo phylum desde los tiempos del Cámbrico Inferior. Bien es cierto también que el mundo no ha estado tan vacío como después de la extinción del Pérmico o antes de la explosión del Cámbrico, pero algunas situaciones locales han surgido desde entonces, como nuevas tierras que emergen del mar o los continentes insulares al ser invadidos por primera vez por nuevos grupos, que aunque no son grandes barriles si son cuencos de un tamaño considerable.
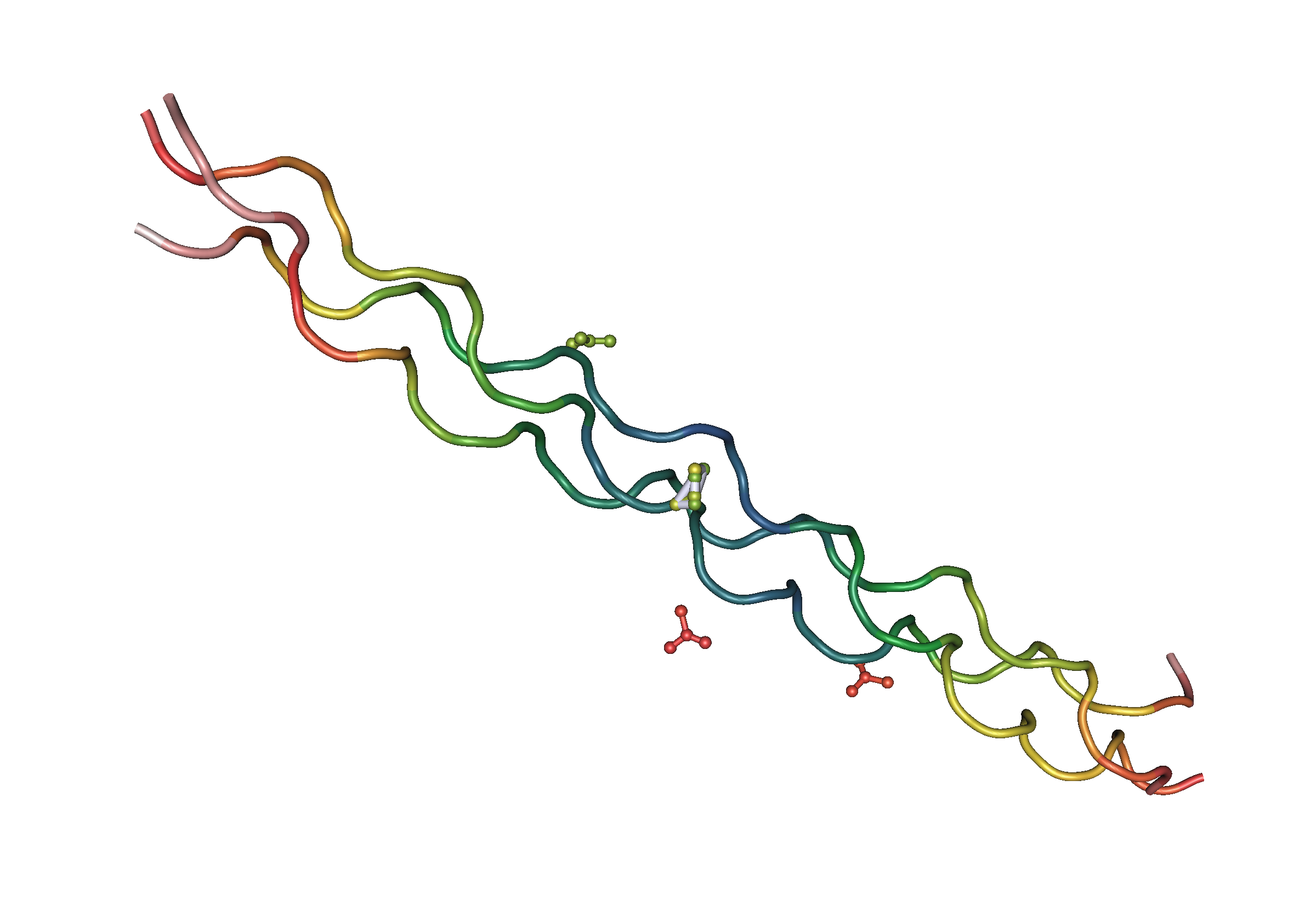
Una representación de la estructura de un colágeno.
Así, se propone como alternativa que las diferencias en las tasas y modelos de cambio evolutivo son respuestas de un sustrato material alterable (los genes) que envejece (en el sentido de que se vuelven, citando a J. W. Valentine“menos dispensadores de reestructuraciones anatómicas importantes”). Así, los organismos modernos no pueden engendrar una rápida colección de diseños nuevos debido a la pérdida de plasticidad para hacerlo, pues según afirman, los genomas de las especies cámbricas serían más flexibles (que no simples). La evolución de múltiples copias para muchos genes, muchos de los cuales codifican para variantes de una misma proteína (como el colágeno) o que divergen después en una gama de funciones relacionadas parece haber atado a los genomas en redes complejas de interacción que no serían ya fáciles de romper debido a su intrincada interrelación. ¿Se desarrollaron los organismo más antiguos con más traducción directa de gen a producto, permitiendo a estos organismos intercambiar y/o alterar partes de su cuerpo separadamente?

Distintas Drosophila melanogaster. El genoma de una especie sería como las piezas de un mecano con las que jugaría la evolución. Estas piezas contienen las instrucciones para hacer una mosca.
Según los seguidores de esta tesis, siempre se ha considerado que las criaturas que no estaban emparentadas de manera profunda y que reaccionaban del mismo modo a un mismo estímulo, por lo general lo hacían asumiendo la existencia de alguna fuerza externa a la genética, pues para los darwinistas clásicos, los sistemas genéticos son demasiado desiguales. Esta fuerza externa sería la única causa común plausible para explicar este problema. Para los seguidores de estos postulados, la semejanza por convergencia evolutiva de dos especies no emparentadas frente a un problema común no se debe a que sea la mejor solución, sino a que la evolución, en su manera de operar no conoce otra manera de solucionar el problema más que con unas instrucciones fijas.
A mi me gusta comparar esta tesis con un juego de mi tierna infancia: el mecano. Así, la evolución por medio de las piezas de este “mecano” genético, antaño dió soluciones más amplias a un mismo problema común, desembocando esto en una mayor disparidad anatómica y funcional. A lo largo del tiempo, esos organismos se extinguieron y es como si al extinguirse, la evolución hubiese perdido una serie de piezas de este particular juego con ellos, por lo que para solventar un problema común entre especies no emparentadas, no le quedará más remedio que hacerlo recurriendo siempre al mismo andamiaje, retocando tan sólo ciertos remates finales a la hora de confeccionarlo.
